
The global prevalence of metabolic syndrome (MetS) has reached alarming proportion with ~30% of the world’s adult population being affected marking a significant public health crisis driven by obesity, sedentary lifestyle, and ageing. Our group is investigating obesity at the molecular level by studying lipid droplets (LDs), an organelle dedicated to fat storage. LDs are found in all cell types, including skeletal muscles, heart muscles, kidney, liver and brain cells. However, in adipocytes LDs are strikingly large, unilocular and they fill the entire volume of the cell. Though, LD biogenesis occurs in virtually all cell types, and an incremental progress has been made in recent years towards an understanding in the molecular mechanisms of this pathway, our knowledge is far from complete. Dysregulated lipid metabolism manifests in various pathological conditions such as obesity, type-2 diabetes, insulin resistance, fatty liver, lipodystrophy, cancer and neuronal diseases. A more detailed understanding of how LDs are formed is thus important to apprehend the etiology of these syndromes.
“Biogenesis of sterol ester-rich lipid droplets employ distinct molecular factors from those involved in triacylglycerol-rich lipid droplet formation”
LD biogenesis occurs in the endoplasmic reticulum (ER), a membranous labyrinth within the cytoplasm where enzymes catalyzing production of storage fats/neutral lipids (NLs), triglycerides (TAG) and sterol (cholesterol) esters (SE) resides. Previously we have demonstrated that LD formation in response to metabolic cues is restricted to few highly defined and specialized ER subdomains demarcated by ER transmembrane protein seipin. Cryo-electron microscopy studies have revealed how seipin assemble into oligomeric complex at LD biogenesis sites. Interestingly, seipin is a non-enzymatic protein defects in which results in lipodystrophic syndrome characterized by selective loss of adipose tissue, along with neurological problems collectively known as seipinopathies. Seipin at LD biogenesis sites collaborates with several LD biogenesis factors to establish the machinery of droplet formation. Seipin performs a decisive role in initiating LD biogenesis, absence of which results in ectopic TAG packaging, resulting in abnormally small or oversized LDs that cannot function properly, thereby affecting overall fat metabolism. However, how these sites are selected and regulated remains largely unknown. Most of the studies by far have focussed on the assembly of TAG-rich LDs, where critical role of seipin has been established, however, the mechanism of biogenesis of SE-rich LDs remains largely unknown. In mammals, steroidogenic tissues such as the testis, ovary, and adrenal glands specialize in storing SE-rich LDs, that undergo rapid degradation upon demand for steroid hormone production. Hence, better understanding of SE-rich LD biogenesis and its turnover will provide molecular insights into flux of sterols for steroid hormone biosynthesis.
We used specially engineered baker’s yeast, Saccharomyces cerevisiae, to investigate the origins of TAG-rich/SE-rich LDs. Since both yeast and humans store excess fats into LDs, yeast serves as an excellent model to elucidate the molecular details of this pathway. Remarkably, most of the findings in yeast is conserved in mammals and mammalian orthologues can functionally complement yeast mutants. For example, human seipin can rescue yeast seipin (Sei1) deletion. We have elegantly shown that seipin defined ER sites exist even in cells that have no LDs, and that upon initiation of either TAG or SE biosynthesis using genetic mutants, cells begin to produce droplets that contain exclusively TAG-rich or SE-rich LDs respectively at the seipin-complex (Figure 1A, B).

Surprisingly, we found that a truncated form of seipin, Fld1-∆LR in yeast, and the human equivalenthSeipin-∆LR, mutants devoid of the conserved luminal domain region (LR) can functionally complement the role of wild-type seipin. Remarkably, truncated seipin containing the transmembrane domains and lacking the LR mimics functional wild-type seipin in defining ER sites that recruits the full LD biogenesis machinery resulting in de novo LDs being born at these locations. Interestingly, we demonstrate that Fld1-∆LR can oligomerize independently of the LR facilitated by protein-protein interaction between Fld1-∆LR monomers. We found that yeast seipin mutants expressing Fld1-∆LR/hSeipin-∆LR had markedly increased steady state TAG levels, indicating important regulatory role of the luminal domain in seipin. Our data suggests that luminal domain in seipin is dispensable in defining functional LD biogenesis sites in the ER, thus providing mechanistic insights into the process of LD assembly.
“The luminal domain region of seipin is dispensable in defining functional sites of lipid droplet biogenesis in the ER.”
More recently, we have shown that similar to TAG-rich LDs, biogenesis of SE-rich LDs also occur at seipin marked ER sites. This has allowed us to investigate this process in greater detail to uncover which proteins are selectively recruited at seipin defined ER sites when cells are actively making TAG-rich vs SE-rich LDs. We provide evidence that several key factors and/or lipid that localize at Sei1 sites during biogenesis of TAG-rich LDs fail to do so when cells are engaged in the production and assembly of SE-rich LDs. We found that deletion of Nem1, Yft2 or Pex30, factors previously known to be crucial for the biogenesis and growth of TAG-rich LDs, were dispensable for the biogenesis of SE-rich LDs. Moreover, neither Pah1, the mammalian lipin homolog, nor diacylglycerol (DAG), a product of Pah1 enzyme and a substrate for TAG synthesizing enzymes accumulated at Sei1 sites upon induction of SE-rich LD formation. This indicates that TAG vs SE production is uncoupled by means of factors that get recruited at seipin sites (Figure 1C). The findings suggest that specialized pathway exist for storage of different class of lipids via recruitment of distinct molecular factors during birth of LD. Identifying novel leads to fine tune this process will have far reaching implications for therapeutic interventions in LD storage disorders.
We further demonstrate that yeast SE acyltransferases, Are1 and Are2, become enriched at seipin defined ER sites during stimulation of SE production indicating that SE synthesis occurs locally, a likely prerequisite for efficient sequestration of SEs and droplet assembly. Consistent with this we have previously shown that TAG producing enzyme, Lro1 gets recruited to seipin sites upon induction of de novo TAG biosynthesis. This indicates that cells have mechanisms in place that control localized accumulation of TAG/SE so as to prevent the detrimental consequences of ectopic NL synthesis throughout the ER membrane. Seipin along with accessory factors would bind to TAG/SE and perhaps act as gatekeepers, thereby allowing growth and maturation of nascent LDs.
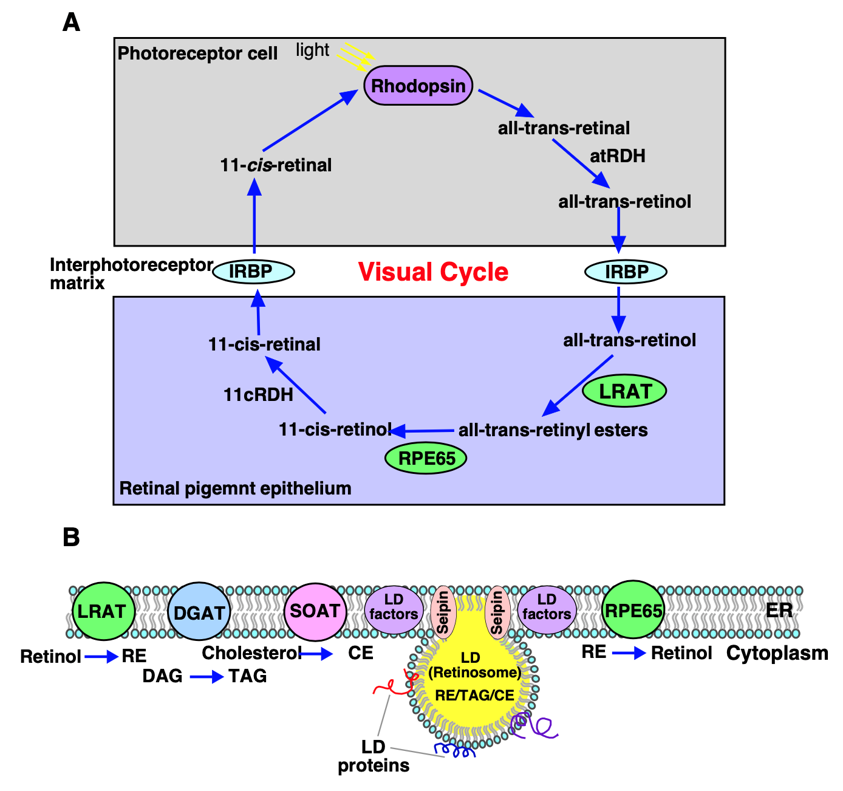
In a recent review we highlighted the importance of lipid homeostasis in the retina to ensure proper visual functioning and how dysregulation in this process due to genetic or acquired factors is implicated in the pathogenesis and progression of several retinal diseases. The visual cycle is a continuous biochemical pathway in the retina between the photoreceptor cells (rods and cones) and the retinal pigment epithelium (RPE) that regenerates 11-cis retinal, a light sensitive molecule essential for vision (Figure 2A). Upon light absorption in the photoreceptor cells, 11-cis retinal undergoes step-wise transformation to all-trans retinol (vitamin A) form that translocates to the RPE cells. In the RPE all-trans retinol is esterified by ER localized enzyme lecithin-retinol acyltransferase (LRAT) to retinyl-esters (RE) that are deposited into morphologically unique lipid droplet (LD) structures called retinosomes (Figure 2A, B). Apart from RE, retinosomes store TAG, SE, and cholesterol. Upon demand REs are hydrolyzed by an ER localized isomerohydrolase, RPE65 generating 11-cis retinol that upon oxidation forms 11-cis retinal that moves back to photoreceptor cells, combines with opsin and rebuilds rhodopsin thereby completing visual cycle (Figure 2A, B). Several mutations in RPE65 have been implicated in retinal pathologies including Leber’s congenital amaurosis (LCA), and retinitis pigmentosa (RP), underscoring the important role of RPE65 in the visual cycle. Hence LDs/retinosomes in the retina play a crucial role in visual chromophore homeostasis for maintaining normal vision. How seipin might promote sequestration and formation of RE-rich LDs and what factors are involved in this process awaits future investigation. In pathological retina, dysregulated lipid metabolism results in extracellular deposition of aggregated lipids and proteins known as “drusen” that slows down trafficking of nutrients and removal of waste materials between RPE and blood vessels resulting in dysfunctional RPE thereby impairing vision.
The most perplexing question is what determines localization of seipin. Whether it is mediated by protein or lipid cues, or a combination of both remains to be determined. Knowing more about how these ER subdomains are formed will pave the way forward in deciphering the mechanism of LD biogenesis. Mature LDs interact with several organelles via membrane contact sites to facilitate bidirectional lipid transfer. Although several tethering factors have been identified, the dynamics of LD-membrane contact association and its release in response to environmental cues is poorly understood. Although our studies provide insights into distinct mechanisms for storage of TAG vs SE containing LD, it remains to be determined how SE packaging is coordinated with TAG filling into LDs. Moreover, whether seipin plays a role in the biogenesis of RE containing LDs/retinosomes remains largely unexplored.
What we have learnt is that LD biogenesis is an intricately regulated process. Absence of LD biogenesis factors, due to genetic mutation or deletion impairs droplet formation, resulting in ectopic NL production and deposition of aberrant LDs, manifesting in ER stress and lipotoxicity, thereby impacting cell physiology and metabolism. Elucidating how this process maintains energy homeostasis, provides key insights into metabolic disorders such as obesity, diabetes, insulin resistance, and lipodystrophy.
“Retinosomes in the retina play a crucial role in visual chromophore homeostasis for maintaining normal vision.”
Moving forward, we would like to characterize this process in mammalian cells to better understand how cells handle sudden influx of fatty acids for efficient storage into LDs in a regulated manner thereby preventing lipotoxicity. It would be interesting to determine how different domains in seipin might be orchestrating an organized filling of TAG/SE into LDs. It remains to be determined if mammalian SE-acyltransferases SOAT1/SOAT2 would also localize at seipin sites for the synthesis of SE-rich LDs. How does the Sei1-complex concentrate SEs/REs? Knowing how seipin interacts with its partner proteins during lipogenesis or lipolysis will likely reveal how sites for LD biogenesis are selected and what role LDs play in energy metabolism homeostasis at cellular, tissue and whole organism’s level. Findings in the coming years will bring novel insights into the pathological conditions that results due to impaired LD biogenesis and/or its defective intracellular trafficking and thus opening new avenues for developing therapeutic leads for lipid storage disorders.
Acknowledgement
This work was supported by an Indo Swiss Joint Research Programme of Department of Biotechnology (Grant: IC12044(11)/6/2021-ICD-DBT awarded to VC), an Extramural Grant of the Indian Council of Medical Research (ICMR), New Delhi (Grant: IIRPSG-2025-01-01895 awarded to VC) and DBT/Wellcome Trust India Alliance Fellowship (GrantIA/I/20/2/505191 awarded to VC). AB is supported by Ramanujan Fellowship of Science and Engineering ResearchBoard (SERB), Department of Science and Technology (DST), Government of India (Grant: RJF/2019/000040), and theCore Research Grant (CRG) of SERB, DST (CRG/2021/003472).










{kind=link}